Fêmea
Fase Folicular
- Inicia-se após luteolise por redução da progesterona que deixa de inibir GnRH.
- GnRH leva à libertação de FSH e LH promovendo desenvolvimento folicular e produção de estrogenio.
- Foliculos antrais desenvolvem-se como resposta a niveis basais de FSH e LH.

GnRH
- Libertado pelo hipotalamo.
- Estimula a libertação de FSH e LH.
- Inibida pela progesterona.
- Centro tónico (nucleo ventromedial e arcuato) libertam pulsos basais.
- Centro ciclico (nucleo preoptico, supraquismatico e hipotalamo anterior) liberta o pico pre-ovulatório de GnRH, que estimula o pico de LH, quando o estrogenio passa um valor limite e a progesterona está baixa. No resto do tempo tem pulsos basais.
- Gerador de pulsos leva a descargas intermitentes. Pode sofrer acção de factores: nutrição, puberdade, stress, lactação, fotoperiodo, ferormonas.
Fases
- Recrutamento: desenvolvimento folicular de um grupo de foliculos antrais por aumento da FSH. Alguns sofrem atresia, outros passam a dominantes. Produzem concentrações crescentes de estradiol e produzem inibina que inibe a libertação de FSH. Em especies monotocos (nasce apenas 1 cria) apenas um foliculo desenvolve dominancia por inibição dos outros foliculos antrais (por inibina e estradiol), reduzida circulação nesses foliculos e FSH em baixas concentrações que resulta em atresia.
- Selecção: selecção de parte dos recrutados e atresia dos restantes, produção de estradiol moderada e inibina (inibe FSH), LH aumenta.
- Dominancia: os selecionados tornam-se dominantes e produzem grandes quantidades de estradiol. Este promove o pico de LH. Como FSH se mantem baixo, os foliculos antrais sofrem atresia. Após o pico pre-ovulatorio de LH, o estradiol cai.
- Atresia: ocorre continuamente durante a foliculogenese (90%).
Ondas
- No metaestro (1ª onda) e diestro (2ª onda) há recrutamento mas devido à elevada progesterona sofrem atresia.
- Após luteolise e queda na progesterona, a 3ª onda de foliculos desenvolve-se dando origem ao foliculo pre-ovulatorio.
- Se o CL regride precocemente por aplicação de PGF2a, a 1ª e 2ª onda podem originar um foliculo dominante pre-ovulatorio.
Sintese de Estrogenio
- A teca interna produz testosterona a partir de colesterol na presença de LH. Difunde a testosterona para as celulas granulosas que a convertem a estradiol na presença de FSH. Os niveis de estrogenio induzem o pico pre-ovulatorio de LH e a produção de receptores de LH nas células granulosas.
- Efeitos do estrogenio: espessamento da mucosa vaginal, aumento das secreções mucosas (cervix, vagina, oviducto), desenvolvimento das glandulas uterinas, aumento da frequencia dos cilios do oviducto, aumento do fluxo sanguineo, leucocitose, edema da genitalia externa, aumento do tónus e motilidade do miometrio, alteração da condutividade electrica, receptividade sexual.
- Papel dos esteroides: manter funcionalidade dos órgãos reprodutivos, regulação das hormonas gonadotroficas, comportamento (sexual, agressão), características sexuais secundarias, influencia metabolismo, crescimento, e desenvolvimento da glândula mamaria.
Ovulação
- Aumento a pressão folicular: teca interna edematosa por aumento da permeabilidade vascular, factores angiogenicos, hiperemia local (PGE2 e histamina).
- Fragilidade da parede folicular: após o pico de LH, a teca interna começa a produzir progesterona que estimula a sintese de colagenase que degrada a tunica albuginea. PGF2a causa ruptura dos lisossomas das celulas granulosas levando a maior degradação do tecido conjuntivo. PGE2 ajuda a remodelação em corpo luteo (activa o plasminogeno).
- Contrações ovaricas: PGF2a causa contracções aumentando a pressão e forçando a protrusão do estigma.
Ovulação induzida
- Ovulação espontanea ocorre em frequencias regulares como resposta a alterações hormonais.
- Ovulação induzida (reflexa) requer estimulo na vagina ou cervix para ovular durante um perido longo ou elevada frequencia (coelhos, gatos, doninhas, camelideos). Há condução do estimulo da vagina/cervix, para a medula espinhal e desta ao hipotalamo que liberta um pico pre-ovulatorio de GnRH. Copulas multiplas induzem picos de LH maiores. Pode-se induzir artificialmente por estimulo mecanico ou electrico.
- Existe um pico pre-ovulatorio de LH induzido pela cópula. Existe também um factor indutor de ovulação (OIF) no plasma seminal (experiências em que ovulam com tratamento de plasma seminal: 90% lamas, >75% camelas asiáticas, e com sémen de touro em 29% dos lamas).
Glândula pineal

- Estrutura cónica constituida por pinealócitos (cels secretoras e cels da glia), no cérebro junto ao hipotalamo.
- Sem barreira hemato-encefálica: fácil penetração por substâncias de peso molecular elevado.
- Recebe “inputs” do sistema nervoso autónomo simpático, não tem contacto directo com o sistema nervoso central.
- Melatonina: principal substância activa produzida pela glândula (indolamina formada do triptofano) que aumenta nos periodos de escuridão e diminui nos periodos de luminosidade (ratinho, homem, égua, ovelha, cabra). A principal função é controlar o desenvolvimento e ciclicidade sazonal das gonadas, estimulando ou inibindo a síntese ou secreção do FSH e LH através da modulação da secreção de GnRH.
Foliculogenese e Ovulação artificial
- Ovulação induzida requer luteolise prematura por PGF2a exogena.
- Superovulação (nº de foliculos anormalmente elevado) requer administração de eCG ou FSH seguida de LH, GnRH ou hCG alguns dias depois. O FSH elevado leva a recrutamento e selecção de mais foliculos.
Maturação do oocito
- Divisão mitotica de celulas germinais primordiais prenatal.
- Nucleo dormente na profase meiotica (dictiotene), até estimulo por gonadotropinas após a puberdade ou ovulação.
- Crescimento citoplasmatico durante o crescimento somático da fêmea. O oocito primordial acumula citoplasma e desenvolve zona pelucida e gap junctions (mediado por celulas granulosas).
- Recomeço da meiose quando o oocito atinge a fase dominante (tamanho minimo critico e há pico LH pre-ovulatorio). Disrupção da comunicação entre oocito e celulas granulosas remove a inibição da meiose (inibidores: AMP, OMI).
- Potencia hormonal: padrão e duração da secreção, semi-vida, densidade de receptores, afinidade hormona-receptor.
- Objectivo do uso de hormonas: maneio reprodutivo (sincronização, ovulação), terapêutica (atonia uterina), diagnóstico.
Factores de Crescimento Semelhantes à Insulina (IGF)
- Produzidos em diversos tecidos, nomeadamente no fígado e no folículo ovárico.
- Papel importante na potenciação do efeito do LH no CL e na oogénese (IGF-I).
- Essencial para o desenvolvimento normal do feto (IGF-II).
Fase Luteica
Formação do corpo luteo (luteinização)
Antes da ovulação a membrana basal degrada-se e permite mistura de células da teca interna e granulosas. Na ovulação, o foliculo implode e há ruptura de vasos sanguineos, formando-se um coagulo (corpo hemorragico) que dura 1 a 3 dias. Por acção do LH, as células da teca interna e granulosas sofrem luteinização em células luteais pequenas (vesiculas de lipidos) e células luteais grandes (granulos de oxitocina e relaxina), respectivamente. Ambas são esteroidogénicas (produzem progesterona). Pode manter-se uma cavidade remanescente do antro folicular. No 3º a 5º dia o corpo luteo (CL) aumenta de tamanho, perde aparencia hemorragica e tem cor laranja (B-carotenos). O aumento de tamanho ocorre por hipertrofia (aumento de volume) das células luteais grandes e hiperplasia (aumento em nº) das células luteais pequenas. Fibroblastos, capilares e eosinofilos tambem aumentam em nº. A vascularização é essencial à sintese e libertação de hormonas. O aumento da massa corresponde à produção máxima de progesterona no diestro. Quando ocorre luteolise, forma-se uma cicatriz (corpo albicans) que perdura por alguns ciclos. Função luteal insuficiente causa falha reprodutiva porque o utero não sustenta o embrião.- Vacas: palpação transrectal permite identificar o CL mas não indica se é funcional. Apenas com ecografia se pode avaliar porque o diametro do CL relaciona-se com a progesterona.
- A progesterona é muito maior nos suínos porque há vários CL.
- Na égua o CL não é palpavel porque está no cortex do ovário.
Progesterona
- Necessario LH basal e colesterol para a sintese.
- Estimula secreção máxima no endometrio glandular - suporta o embrião.
- Inibe o miometrio reduzindo contracções - permite implantação.
- Estimula desenvolvimento da glândula mamaria antes do parto.
- Feed-back negativo no hipotalamo (diminui frequencia basal de GnRH). No entanto o LH está elevado e o FSH tónico, permitindo o desenvolvimento de foliculos mas sofrem atresia.
- Progesterona elevada impede formação de foliculos pre-ovulatorios, produção de estrogenio, estro e pico pre-ovulatorio de GnRH e LH.
- Sensibiliza o cerebro aos efeitos do estrogenio.
Luteolise
- Degeneração do CL com queda na progesterona. Se a luteolise não ocorrer mantem-se em fase lutea.
- PGF2a: o aumento da progesterona é acompanhado por diminuição dos seus receptores no endometrio. O utero ipsilateral exposto a progesterona elevada por um determinado tempo começa a produzir PGF2a. Aos 10 a 12 dias a progesterona deixa de conseguir bloquear os receptores de oxitocina no utero (oxitocina luteal), que causa maior libertação de PGF2a (feed-back positivo). O PGF2a é libertado na veia utero-ovarica que cruza em contracorrente com a arteria ovárica, evitando dilução (PGF2a desnaturada na ciruclação pulmonar). Na fase luteica tardia ocorrem pulsos de PGF2a que devem atingir uma certa frequencia para induzir luteolise completa.
- No cão o utero ipsilateral não tem muita influencia. Na égua não há degradação na circulação pulmonar e o CL é mais sensivel.
- PGF2a exogena após o 6º dia causa regressão do corpo luteo.
- Degeneração capilar: reduz a vascularização do CL.
- Apoptose: macrofagos e linfocitos no CL produzem citocinas que inibem a sintese de progesterona e activam a apoptose. De seguida ocorre limpeza e apenas fica tecido conjuntivo que forma o corpo albicans.
Sincronização de estros
- Progesterona exogena: suprime o estro e ovulação. Quando removida entra em proestro e estro em 2 a 3 dias. Permite sincronizar cios para IA.
- Progesterona Intravaginal (CIDR)
- Sincronização em bovinos.
- Administrado por 7 dias com administraççao de PGF2a (Lutalyse) ao 6º dia.
- A progesterona impede libertação de GnRH e gonadotropinas, desenvolvimento folicular e ovulação. A PGF2a inicia luteolise.
- Ao remover o CIDR há uma quebra na progesterona que induz pico de GnRH, o que causa pro-estro e estro em 2 a 3 dias e insemina-se.
- Administra-se novamente o CIDR apás 14 dias da IA mas sem PGF2a. Animais gestantes não regressam ao cio.
- PGF2a: luteolisina que encurta o ciclo estrico e induz cio em bovinos quando administrada entre o 7º e 18º dia (o CL da vaca não é sensivel nos primeiros dias).
- Ovosync
- Permite sincronização para IA sem necessidade de detecção de cio.
- 1º: GnRH - se há foliculo dominante (>10mm) ovula, se não há foliculo dominante promove o crescimento de foliculos (ainda tem CL da ovulação anterior).
- 2º: PGF2a após 7 dias - causa luteolise e entrada na fase folicular.
- 3º: GnRH após 48h - induz ovulação.
- 4º: IA após 16h, sem necessitar de detecção de cio.
- Problemas: GnRH não induz ciclicidade se em anestro, ovulação com menor concepção, podem recrutar foliculas da 2º e 3º onda que não ovulam, PGF2a não é totalmente eficiente. Solução: Pre-sync.
- Pre-sync
- Começa 26 dias antes do primeiro GnRH, administra-se PGF2a que faz lise do CL se existente.
- Passado 14 dias administra-se PGF2a que causa lise do CL nas vacas que anteriormente estavam em fase folicular.
- Passado 12 dias GnRH que causa ovulação do novo foliculo formando um novo CL.
Macho
Regulação endocrina
- O macho não tem centro de ciclos.
- GnRH é libertado intermitentemente, causando pulsos de LH essenciais à produção de testosterona (se LH sempre alto, as células de Leydig tornam-se refractarias - reduzem o nº de receptores e diminuem a secreção de testosterona).
- FSH tem pulsos mais baixos e longos (produção de inibina nos testiculos).
- Celulas de leydig (~ teca interna): sofrem acção da LH, e produzem progesterona que é convertida em testosterona (libertação pulsatil e curta - 4 a 8 picos em 24h). A testosterona intratesticular é 100 a 500x mais elevada porque tem pequena semi-vida, é diluida na circulação sistemica.
- Células de Sertoli (~ cels granulosa): dependem do FSH, convertem testosterona a estradiol e dihidrotestosterona. O estradiol faz feed-back negativo no hipotalamo (GnRH). Tambem produz inibina que inibe a libertação de FSH.
Espermatogenese
- Nos tubulos seminiferos, durante o desenvolvimento movem da membrana basal para o lumen. As células germinais estão ligadas por pontes intercelulas que comunicam. A produção é continua excepto em reprodutores sazonais.
- Proliferação: a espermatogonia (2N) na membrana basal sofre divisões mitoticas resultando em espermatogonias-B. Há três tipo de espermatogonia (A1, A2, A3, A4, I, B). Stem cells dividem-se mitoticamente para renovar espermatogonias A.
- Meiose: a divisão mitotica de espermatogonia-B forma o espermatocito primário que entra em profase meiotica (fase que ocupa 30% do tempo). Há crossing-over do DNA. Forma-se um espermatocito secundario que imediatamente sofre a segunda divisão meiotica em espermatideos (1N).
- Diferenciação (espermiogenese): transformação do espermatideo em espermatozoide.
- Fase de Golgi: o complexo de Golgi bem desenvolvido junto ao nucleo produz vesiculas pro-acrossomicas que se fundem num polo do nucleo e formam a vesicula acrossomica (contem um granulo acrossomico). Os centriolos movem-se para a base do nucleo, dando o proximal orgem ao complexo de implantação (ancora o flagelo ao nucleo) e o distal o axonema (porção central do flagelo).
- Fase da capa: o acrossoma forma uma capa no nucleo anterior. O complexo de golgi move-se caudalmente e desaparece. O flagelo primitivo começa a projectar-se.
- Fase acrossomica: o acrossoma expande-se ate cobrir ⅔ do nucleo anterior. O nucleo alonga-se e os microtubulos (manchette) ligam-se ao nucleo possterior ao acrossoma formando a capa pós-nuclear. Os espermatideos estão nas células de Sertoli com caudas a protrudir.
- Fase de maturação: a manchette migra para a cauda e desaparece ficando só parte a formar a capa pó-nuclear. As mitocondrias migram e formam uma espirar à volta do flagelo formando a peça intermedia. Formam-se as fibras exteriores densas e a bainha fibrosa do flagelo.
- Espermiação: libertação do espermatozoide para o lumen. A cabeça do espermatozoide (caracteristica da espécie) é longa contendo um nucleo oval com cromatina compact e queratinizado. O acrossoma contem enzimas hidroliticas (acrosina, hialuronidase, lisina da zona, esterase, hidrolases acidas) que permitem penetrar a zona pellucida do oocito. A cauda tem o capitulo (permite implantanção) e colunas laminadas que dão flexibilidade. É contituido por 2 filamentos centrais rodeados de 9 pares de microtubulos, e o grupo rodeado de 9 fibras mais grosseiras. As mitocondrias em helix formam a peça intermedia. O anulo permite junção entre a peça intermedia e a cauda.
Caracteristicas
- Ciclos do epitelio seminifero: considera-se que a progressão do ciclo 1 ao 8 demora 13,5 dias. Uma espermatogenese de 61 dias (touro) tem 4,5 ciclos.
- Produção de esperma diaria: nº total de espermatozoides produzidos por ambos os testiculos. Necessita de retirar uma amostra de parenquima. O nº por dia por grama é referido como eficiencia de produção de esperma e depende no nº de celulas de Sertoli.
- A capacidade de produção de esperma pode ser avaliado colhendo esperma ou medindo a circunferencia de ambos os testiculos (maior, maior produção).
- Motilidade: é a capacidade de movimento linear progressivo (% estimada) a 37º (motilidade pára a 18º), relaciona-se com a viabilidade. Avalição da viabilidade pode ser feita por consumo de oxigenio, colorações e citometria de fluxo.
- Anormalidades: o aceitavel é entre 5 a 15%, ocorre por erros na diferenciação, transito ou maturação. Algumas são hereditárias. Outros erros não visiveis são erros de função (composição nuclear ou bioquimica anormal, defeciencia em proteinas de superficie, deficiente resposta aos estimulos).
Recolha de sémen
- Após colheita avaliar: volume, motilidade progressiva (> 60%), concentração. A concentração multiplicado pelo volume dá o numero total de espermatozoides no ejaculado. Para determinar o nº de doses para inseminação, divide-se o nº total de espermatozoides pelo nº para cada dose.
- Extenders: isotonico, tampão (muito alcalino ou acido causam morte celular; tris, citrato de sodio, fosfato de sódio), crio-protector (protege entre 0 a -50; penetrante são DMSO e glicerol; não-penetrantes são proteinas do leite e gema de ovo), nutrientes (espermatozoides são dependente do ambiente; frutose e glucose) , evitar crescimento microbiano (penicilina, linquamicina, linco-spectina, estreptomicina), manter viabilidade, low-cost.
- Separação de sexo: o cromossoma X tem mais 2.8 a 4.2% de DNA e é mais corado que o cromossoma Y, por citometria de fluxo, o X emite luz mais intensa e permite separação por citometria de fluxo (caro, lento).
Esperma no trato genital da fêmea
 |
| In Pathways of Pregnancy and Parturition, 2nd ed, Senger |
Transporte
- Fase rápida: por contracção da musculatura da fêmea, função desconhecida.
- Fase continua: uniforme num longo periodo de tempo, transporte de locais de reserva do cervix e junção uterotubal.
- O estradiol e prostaglandinas do esperma (PGF2a e PGFE1) estimulam contracção do mimetrio impulsionando o esperma cranial e caudalmente. O fluido serve de veiculo.
- O cervix tem sulfomucinas (muco viscoso) na região apical e sialomucinas (muco fluido) na região basal das criptas. Os espermatozoides que nadam pelas criptas com sialomucina têm uma via priveligiada. O cervix filtra espermatozoides não moveis.
- Na vaca, o sémen depositado num corno tem transporte para o outro e a colocação no cervix têm maiores perdas.
Capacitação
- O esperma tem moleculas de membrana que são cobertas por proteinas do plasma seminal. Na capacitação há perda das cobertura seminal e parte das moleculas de superficie (permite penetral e fertilizar o oocito). Inicia-se no utero e completa-se no oviducto.
- Aumento da permeabilidade da membrana ao calcio. Modificação da membrana celular (remoção de glicoproteinas). Libertação da hialuronidase (penetração cela. granulosa) e conversão de pro-acrosina a acrosina (penetração ZP).
- Secreções facilitam capacitação e reacção acrossomica: aminoacidos, proteínas, frutos, galactose, acido sialico e citrico, vitaminas, factores de crescimento. Glicoproteinas especificas do oviducto (EGP) são particularmente importantes e são sintetizadas em resposta a um aumento dos estrogénios entre os dias -1 e 3 (dia 0 = estro).
Fertilização
- Na ampola do oviducto, por contacto com proteinas especificas torna os espermatozoides hiperactivos (nadam em circulos em pequena área) que facilita o contacto com o oocito.
- As proteinas especificas da membrana do espermatozoide (expostas na capacitação) ligam-se à proteina ZP3 da zona pelucida. A zona de ligação primaria permite aderencia enquanto o ligando promotor da reacção acrossomica inicia a reacção acrossomica.
- A reacção acrossomica inicia-se com a fusão da membrana exterior do acrossoma com a membrana celular formando pequenas vesiculas (vesiculação) que libertam enzimas (ex. acrosina). Quando terminada o nucleo fica com a membrana interior do acrossoma. A força do flagelo empurra o espermatozoide pela zona. Tem como objectivo penetrar a zona pelucida e expor o segmento equatorial. A zona fica predominantemente intacta, importante porque impede dispersão de celulas do blastomero.
- No espaço perivitelino, o oocito forma microvilosidades que suportam o espermatozoide. A membrana começa a fundir-se com o segmento equatorial (proteina de fusão). Após fusão, o oocito sofre uma reacção cortical onde faz exocitose de granulos corticais que bloqueiam a zona e impedem a penetração por outros espermatozoides impedindo polispermia (reacção de zona). A membrana do oocito (oolema) perde qualidade de fusão com novos espermatozoides (bloqueio vitelino).
- O espermatozoide no citoplasma do ovo forma o pronucleo masculino após hidrolise das ligações disulfidicas (queratinizado) do nucleo por glutationa (11 a 39 h após ovulação na vaca; 3 a 9 h na ovelha). Os pronucleos masculino e feminino fundem (singamia) e o zigoto entra nos primeiros estagios de embriogenese.Em especies multiparas, espermatozoides de vários machos podem fertilizar oocitos (superfecundação).
Desenvolvimento
- O zigoto sofre divisões por clivagem originando o blastomero e morula. Se o blastomero se separar pode formar gêmeos.
- As celulas da porção interna forma gap junctions e origina o embrião, a porção externa tem tight junction e forma o trofoblasto. A bomba de Na+ activa permite acumular liquido dentro do embrião e forma o blastocelo (quando reconhecivel chama-se blastocisto).
- A acumulação de liquido com crescimento, proteinas enzimaticas, enzimas proteoliticas e contracções do blastocisto levam à ruptura da zona pelucida. É completamente dependente do ambiente uterino (função luteica e progesterona).
- No periodo pre-ligação formam-se as membranas extra-embrionarias. O blastocisto livre cresce e forma-se a endoderme que reveste o trofoblasto e tambem forma a evaginação (saco vitelino, contribui com celulas para a região genital). O trofoblasto e mesodermo forma o corion que envolve o embrião, formando também o amnio que proteje o embrião de choques e é anti-adesivo para células. Evaginação da região posterior forma a alantoide (recolhe produtos de excreção) e quando entra em contacto com o corion forma o alantocorrion, parte fetal da placenta.
Blastogenese
- Inicio na fertilização, fim na eclosão.
- Duração: <1 semana (ratinho, rato, coelho) 1 semana (vaca, égua) 2 semanas (outras espécies).
- Nutrição: leite uterino/histotrofo (proteínas e lípidos), reservas citoplasmáticas.
- Zigoto/embrião dividem-se uma vez/dia.
- Embrião entra no útero 3 a 5 dias após a fertilização, com 16 células (blastómeros). Excepções:

- Oócitos não fertilizados também são transportados para o útero (exc. Égua)
- Durante a blastogénese tolera períodos de exposição a condições extra- corporais (transferência de embriões).
Embriogenese
- Início na eclosão, final na implantação (mal definido, formação de aderencias e contacto hematogenico entre trofoblasto e endotelio uterino).
- Duração da implantação:

- Diferenciação celular e a organogénese.
- Maior incidência de mortalidade embrionária: 15% em gatas, cadelas e éguas; 30% em vacas, pequenos ruminantes e suínos.
- Período de maior vulnerabilidade a agentes teratogénicos.
Reconhecimento Maternal de Gestação
- O embrião tem que sinalizar a sua presença para impedir luteolise e quebra de progesterona. A oxitocina do endometrio sintetiza PGF2a que origina luteolise e deve ser impedido
- Na ovelha e vaca, há produção de interferões (IFN-τ) pelas células do trofoblasto que impedem a produção de receptores de oxitocina (impede PGF2a) e em parte de estrogenios, e estimulam a produção de proteinas pelas glândulas uterinas (nutrição).
- Na porca, o embrião produz estradiol que redirecciona a excreção de PGF2a para o lumen (incapaz de causar luteolise) e aumenta os receptores de prolactina no endometrio. No entanto é necessario 2 conceptos em cada corno para manutenção da gestação.
- Na égua, o concepto migra (12x por dia) distribuindo proteinas ao longo do utero e inibindo PGF2a (até dia 12 a 18).
- Na cadela e gata, o CL têm duração longa e só sofre luteolise após parto.
Transferencia de embriões
- Sincronização de estadio do receptor e dador. Superovulação do dador (FSH) e inseminação artificial. Recolha por lavagem (tambem por aspiração de oocitos, viaveis 12h). Transferencia para receptor (deposição no corno uterino ipsilateral ao CL).
- Manutenção de oocitos in vitro com temperatura e atmosfera apropriada (5% CO2, 5-8% O2), pH um pouco superior a neutro, ausencia de microrganismos, ambiente ionico e nutricional.
Placentas
- Corion (fetal) + endometrio (maternal) = placenta. Para trocas metabolicas e produção de hormonas.
- Classificação por villi corionicos
- Placenta difusa: villi na superficie inteira do corion (porco), na égua há microzonas com microcotiledones e produz eCG.
- Placenta zonaria: zona central de trocas, anel pigmentado com hematoma (transporte de ferro, paraplacenta), zona distal com poucos vasos (gato, cão).
- Placenta discoide: em dois discos (primata, roedores).
- Placenta cotiledonaria: o placentoma é constituido por cotiledone fetal (corion) e maternal (carunculas) - vacas.
- Classificação histologica
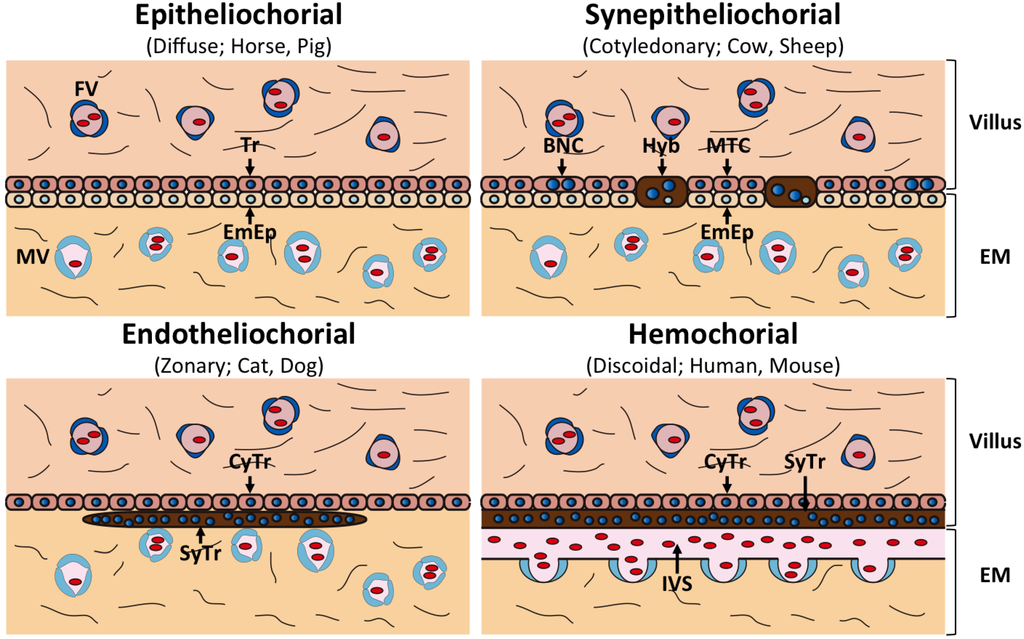
- Epiteliocorial: menos intima, intacta (porca, égua).
- Sindesmocorial: como epiteliocorial com erosões intermitentes, células gigantes binucleadas produzem lactogenio, PSPB, progesterona e estrogenio (ruminantes).
- Endoteliocorial: erosão compelta do epitelio do endotelio, mais intima (cão, gato).
- Hemocorial: aposição directa de sangue (primatas, roedores).
- Função hormonal
- eCG produzido pelos calices endometriais da égua permite manutenção do CL e CL acessorios (originarios de novas ovulações ou luteinização de foliculos antrais por eCG).
- hCG nos primatas promove CL gestacional e produção de progesterona.
- A progesterona é essencial porque estimula secreção das glandulas endometriais e impede as contracções do miometrico. O CL produz no inicio da gestação e pode ser substituido pela placenta (ovelha, egua, human). Os estrogenios produzidos pela placenta são indicadores do periodo pre-parto.
- Somatotropina (lactogenio placental) produzida pela placenta (roedores, ovelhas, vaca, humano) estimula o crescimento do feto e da glandula mamaria.
Parto
Estagio I: inicio das contracções
- A limitação de espaço no utero devido ao crescimento origina stress no feto. Este liberta ACTH que estimula a produção de corticoides, iniciando a cascata de eventos.
- Remoção da acção da progesterona: cortisol promove conversão a estradiol, a placenta sintetiza PGF2a que causa luteolise. Há aumento das secreções.
- A pressão do utero aumenta, o feto coloca-se em posição (ovelha, bovino, equino). O estradiol e PGF2a estimulam contracções do utero. As contracções empurram o feto em direcção ao cervix. A pressão no cervix activa neuronio sensitivos que estimulam a libertação de oxitocina no hipotalamo e esta facilita as contracções. O feto entra no canal de parto.
Estagio II: expulsão do feto
- A relaxina produzida pelo CL ou placenta por estimulaçao por PGF2a permite amolecimento do tecido conjuntivo do cervix e elasticidade dos ligamentos pelvicos.
- O estradiol elevado aumenta a actividade secretoria (especialmente do cervix) reduzindo a fricção e permitindo a saida do feto. Com a pressão, as membranas rupturam e o fluido amniotico e alantoico lubrifica o canal de parto. O feto entra no canal de parto e a hipoxia estimula o seu movimento, que por sua vez estimula contracção do miometrio e dos musculos abdominais (feedback positivo).
- A duração do parto é variavel com a especie. A distocia ocorre principalmente no estagio 2 e deve-se ao tamanho excessivo do feto, falha de rotação do feto ou presença de gemeos.
Estagio III: expulsão de membranas fetais
- Normalmente ocorre rapidamente após expulsão do feto. Os villi desalojam-se das criptas por vasoconstrição.




Muito Obrigada !!
ResponderEliminar